This post was co-authored with Arthur Hunt of the University of Kentucky, who first pointed out these errors on the Peaceful Science forum. I wish I had spotted these myself, but I took Behe’s word on the polar bears because it all sounded solid. This is an important reminder to ALWAYS check the references for yourself. Lesson learned and kudos to Art for catching this and for working with me on this post. -NHL
 The release of Michael J. Behe’s newest book, Darwin Devolves: The New Science About DNA That Challenges Evolution, is nearly upon us. (See my review in Science). The first chapter was made publicly available to entice readers. In this chapter, Behe outlines his main thesis: at the molecular level, adaptive changes are largely due to events that in some way destroy or damage proteins and enzymes. He calls it the first rule of adaptive evolution and to illustrate his point, he discusses the evolution of polar bears and describes the molecular events in that evolution as little more than a series of damaging mutations that result in a more adapted organism.
The release of Michael J. Behe’s newest book, Darwin Devolves: The New Science About DNA That Challenges Evolution, is nearly upon us. (See my review in Science). The first chapter was made publicly available to entice readers. In this chapter, Behe outlines his main thesis: at the molecular level, adaptive changes are largely due to events that in some way destroy or damage proteins and enzymes. He calls it the first rule of adaptive evolution and to illustrate his point, he discusses the evolution of polar bears and describes the molecular events in that evolution as little more than a series of damaging mutations that result in a more adapted organism.
But first, a quick introduction to Behe for those who may not know who Behe is or where this is coming from. With the release of his first book, Darwin’s Black Box, in 1996, Behe helped revolutionize and reorganize the resistance to modern evolution under the banner known as “Intelligent Design,” often abbreviated as ID. Many consider ID as simply creationism by another name, but the ID community works hard to distance themselves from that label. They insist that ID is a scientific theory, not a religious one, based on what they consider evidence that cells and organisms were designed intentionally, rather than the result of the aimless and unguided forces of evolution. Scientists and federal courts disagree, but this has not stopped the steady steamroller of books and articles from the ID community.
Fast forward to 2019. In Darwin Devolves, Behe makes the argument that natural selection, which he prefers to call “Darwinism,” is driven largely, even exclusively, by mutations that degrade or destroy protein function. At the outset, it must be said that we have known for many decades that, occasionally, harming or even destroying a gene or protein can actually be good for the organism. What Behe is saying is that harming genes is pretty much the only way that unguided mutations can ever help an organism. That’s just not the case, but I’ll get to that later.

Back to the polar bears. Behe offers them as an example of how harming genes can help an organism and lead to adaptive evolution. Imagine an ancestor bear population that looked pretty much like brown bears. Then came some random mutations that reduced the production or deposition of pigment into the fur of the bears. This made the bears white and – voilà! – the bears acquired natural camouflage in snowy climates so as to better sneak up on their prey.
This seems like a logical example. Behe uses this example to bolster his claim that this is basically all that unguided mutations can do. However, even in this apparently “pro-Darwinism” example, Behe both exaggerates and misrepresents what science has actually revealed. The evolution of polar bears was not only a matter of harmful mutations, as a close look clearly reveals.
The key reference here is a 2014 paper in Cell. In this study, researchers did genome sequencing of 89 polar bears and brown bears and discovered the molecular changes that distinguish these very closely related species, using the giant panda as the reference sample. The results were fascinating and revealed that polar bears and brown bears have been separate populations with limited gene flow for less than 500,000 years.
 The researchers identified a list of protein-coding polar bear genes that had experienced recent positive selection, meaning that evolutionary forces had strongly favored specific variants. Brown bears, on the other hand, have experienced much less positive selection since the populations diverged. This means that polar bears have experienced stronger selective pressures and have diverged from the ancestors more than brown bears have. This matches what the fossil evidence suggests. Basically, a population of brown bears ventured northward and, in response to the very different climate, evolved into polar bears. The ancestral population stayed pretty much the same and are the brown bears of today.
The researchers identified a list of protein-coding polar bear genes that had experienced recent positive selection, meaning that evolutionary forces had strongly favored specific variants. Brown bears, on the other hand, have experienced much less positive selection since the populations diverged. This means that polar bears have experienced stronger selective pressures and have diverged from the ancestors more than brown bears have. This matches what the fossil evidence suggests. Basically, a population of brown bears ventured northward and, in response to the very different climate, evolved into polar bears. The ancestral population stayed pretty much the same and are the brown bears of today.
One of the genes that experienced the strongest selection is APOB, which encodes apolipoprotein B, a protein involved in the transport of fat molecules in the blood. This makes sense because polar bears subsist on a diet extremely rich in saturated fats, yet don’t develop heart disease with great frequency. Quoting Behe:
The polar bear’s most strongly-selected mutations – and thus most important for its survival – occurred in a gene dubbed APOB, which is involved in fat metabolism in mammals, including humans. That itself is not surprising, since the diet of polar bears contains a very large proportion of fat (much higher than in the diet of brown bears) from seal blubber, so we might expect metabolic changes were needed to accommodate it.
First of all, as shown in Table 1 of the paper, APOB harbors the second most strongly-selected set of variants, not the first, but we can let that one slide.

Behe continues:
But what precisely did the changes in polar bear APOB do to it compared to that of other mammals? When the same gene is mutated in humans and mice, studies show it frequently leads to high levels of cholesterol and heart disease. The scientists who studied the polar bear’s genome detected multiple mutations in APOB. Since few experiments can be done with grumpy polar bears, they analyzed the changes by computer. They determined that the mutations were very likely to be damaging -that is, likely to degrade or destroy the function of the protein the gene codes for.
Some of them, possibly. Definitely not all of them or even most of them, as I’ll soon explain. He continues:
A second highly-selected gene, LYST, is associated with pigmentation, and changes in it are probably responsible for the blanching of the ancestors’ brown fur. Computer analysis of the multiple mutations of the gene showed that they too were almost certainly damaging to its function. In fact, of all the mutations in the seventeen genes that were most selected, about half were predicted to damage the function of the coded proteins. Furthermore, since most altered genes bore several mutations, only three to six (depending on the method of estimation) out of seventeen genes were free of degrading changes. Put differently, 65-83 percent of helpful, positively selected genes are estimated to have suffered at least one damaging mutation.
Now it’s getting harder to excuse Behe’s exaggeration. Specifically, using one specific predictive algorithm, the authors found that only 7 of the 17 genes with the strongest signatures for positive selection are unequivocally predicted to possess at least one “damaging” mutation. Even Behe’s “about half” is just 41%, which means that the lower limit on Behe’s estimation is also wrong. It’s not 65-83%, it’s 41-83%. The range is so wide because computational predictions invariably involve uncertainty.
But more importantly, by Behe’s math, if a gene harbors five enhancing mutations and one diminishing one (by computer prediction), it counts as “damaged.” Any number of gain-of-function mutations can be overtaken by a single damaging one. While that could be true in some cases, a nonsense mutation for example, there is no reason to assume it must be true in all cases.
Considering how little is known about the molecular biology of polar bears, it is entirely possible that none of the 17 most positively selected genes in polar bears are “damaged.” Quoting from the Supplemental information in the paper:
We find no fixed missense mutations specific to the polar bear lineage associated with human diseases according in the Human Gene Mutation Database. However, the top 20 genes are significantly enriched with genes previously associated with metabolic diseases and traits and humans (p-value = 0.042) from the GWAS catalog, discussed in the main text.
In other words, many of the 20 most positively selected genes in the polar bear genome are orthologs of genes that have variants (mutations) in the human population associated with metabolic disease. When these genes are damaged by mutations in humans, the humans are more likely to suffer metabolic diseases. But Behe believes – with no hard evidence – that these same genes, when damaged, protect the polar bears from metabolic disease.
There’s more. If we come back to APOB, the polar bear gene that Behe spends the most time discussing, we find that the authors of the study have a very different interpretation of the data than Behe does. Quoting the paper again [emphasis added]:
Substantial work has been done on the functional significance of APOB mutations in other mammals. In humans and mice, genetic APOB variants associated with increased levels of apoB are also associated with unusually high plasma concentrations of cholesterol and LDL, which in turn contribute to hypercholesterolemia and heart disease in humans (Benn, 2009; Hegele, 2009). In contrast with brown bear, which has no fixed APOB mutations compared to the giant panda genome, we find nine fixed missense mutations in the polar bear (Figure 5A). Five of the nine cluster within the N-terminal ba1 domain of the APOB gene, although the region comprises only 22% of the protein (binomial test p value = 0.029). This domain encodes the surface region and contains the majority of functional domains for lipid transport. We suggest that the shift to a diet consisting predominantly of fatty acids in polar bears induced adaptive changes in APOB, which enabled the species to cope with high fatty acid intake by contributing to the effective clearance of cholesterol from the blood.
Clearly, the authors do not expect the polar bear APOB to be broken or damaged. Rather, a bare majority of the amino acid changes are clustered in the most important region for the clearing of cholesterol from the blood. This argues that these mutations likely enhance the function of apoB, at least when it comes to surviving on a diet high in saturated fats.
It is also worth noting that apoB does much more than clear fatty acids from the blood. It is a very large protein that has many biochemical activities and is a central player for lipid and cholesterol transport. Even if “damaging” mutations might be beneficial in one context, they could very well be harmful or lethal in another. Moreover, mice that lack apoB are not viable.
To recap: 1.) There is no evidence for Behe’s claim that APOB is degraded or diminished in polar bears and everything we know about the protein from other mammals suggests the opposite. And 2.) Behe’s claim that the most common adaptive changes in polar bears are those that degrade or destroy proteins is not supported, and the evidence suggests otherwise. And yet Behe makes this bold claim:
It seems, then, that the magnificent Ursus maritimus has adjusted to its harsh environment mainly by degrading genes its ancestors already possessed. Despite its impressive abilities, rather than evolving, it has adapted predominantly by devolving. What that portends for our conception of evolution is the principal topic of this book.
The word “devolving” actually doesn’t make any sense (here’s why), but the much more important point here is that, the analysis of the polar genome does not support Behe’s claim that the evolution of polar bears was mostly driven by damaging mutations. In fact, the polar bear variant of APOB is almost certainly enhanced compared to the ancestral version, at least in the ways that are important to the challenges of being a polar bear.
<<<<<<<<
IMPORTANT UPDATE: After this post went live, Behe wrote this incredibly uncharitable response in which he called me “incompetent” (and Art Hunt is “some other guy”) and described our review in Science as “mind-bogglingly shoddy.” He implores Jerry Coyne, who had blogged about this post, to “find a more reliable informant” than me. I was pretty surprised at the nastiness in this response, given that Behe has a reputation as being kind and soft-spoken. In fairness, few of us are at our best when caught in an obvious mistake. I digress.
In his response, Behe claims that I misrepresented his position and that he never says that damaging things is all that unguided adaptive mutations can ever do, just that this is mostly what they do. This sounds like a backpedal to me, but I’m perfectly happy to let him speak for himself on how he characterizes the preponderance of evidence. (Richard Lenski, one of my co-authors on the Science review, has written an eloquent blog post about damaging versus innovative mutations.) But also, as you can see above, I block-quoted Behe extensively to avoid any possible charge of “mischaracterizing” his position. He made that charge anyway.
Behe devotes only two sentences to a defense of his position on polar bears and here they are:
Below is the relevant information from Liu et al.’s Table S7. Those who can understand the table will see that it supports every actual, undistorted claim I made about the polar bear.
Behe then pastes this as “the relevant information” from the Table:

^ This ^ is not the original chart that appears in the paper. Behe left out most of the mutations from the original chart. He also left out the results of a different predictive algorithm for characterizing the mutations. Here is the actual Table S7:

I first want to call attention to the two right-most columns of this chart. I have zoomed in on the top half of this chart, to make it easier to inspect:

The two columns that Behe cut off contain the results of a different algorithm for predicting the effect of the mutations. You will notice that this algorithm makes different predictions for many of the mutations, describing many more of them as “benign” and many more of them “possibly” rather than “probably” damaging.
Behe also chose to include only some of the mutations from this Table. I’ve circled all of the mutations that he left out, again focusing only on the top half of the chart:
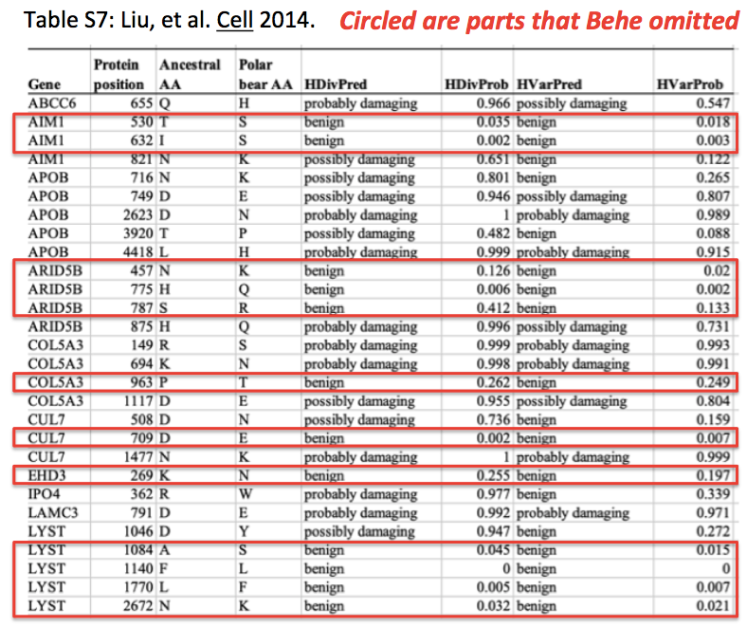
Once again, Behe’s selective presentation of the chart leaves out all of the alterations that are not predicted to damage the protein in which they occur. Combining both of Behe’s omissions into one visual:

In Behe’s defense, he doesn’t explicitly say that he’s presenting the whole Table. So he isn’t lying exactly. Instead, he says that he is presenting “the relevant information” from the Table. I find this deeply misleading. This whole discussion is about the nature of adaptive mutations in the evolution of species and Behe’s arguments is that most of them are damaging. By selectively presenting only the mutations that are predicted to fit his argument, he is intentionally leaving out evidence that is contrary to his position.
After all, what is the purpose of showing the chart at all? To show that some mutations that drove polar bear evolution are damaging? He didn’t need a chart to make that point and no one would argue with that. I suspect that if the unaltered Table S7 gave the impression that the overwhelming number of adaptive mutations were damaging, Behe would have shown the whole thing.
In reality, Table S7 does not give that impression at all, and so he slices it up with surgical precision so that he can present “the relevant information,” that is, the information that appears to support his position. And, at least when it comes to APOB, even the selectively edited information probably doesn’t support his position either, regardless of what the predictive algorithm says, as I (and the study’s authors!) explain above.
The evolution of polar bears is the opening story of Behe’s book, the example he uses to describe his concept of “devolution.” But if you actually consult the data itself, it tells a very different story than Behe does.
<<<<<<<<<
For more of the problems with Darwin Devolves, see my review in Science (co-authored with Richard Lenski and Josh Swamidass, open-access version). I also published a more comprehensive look at the book for Skeptic magazine (and they gave it the cover!).
-NHL (with special guest, Art Hunt)
![]()

[q]They insist that ID is a scientific theory, not a religious one, based on what they consider evidence that cells and organisms were designed intentionally, rather than the result of the aimless and unguided forces of evolution.[/q] It would be interesting to ask, intentionally designed by whom! By “Engineers” from Ridley Scots “Prometheus?” It is absurd. Who designed the “intelligent designers” then?
LikeLike
You don’t need to know who did it before you can determine it was intelligently designed. You don’t even ask about the who or how until after design has been determined. And it stands that to refute any given design inference all one has to do is step up and show that nature could do it.
LikeLike
The cardinal rules of studying antievolutionists (that I try to follow in my own TIP research): don’t take their word for ANYTHING, and FOLLOW THE SOURCES. It will be at that documentation level that the measure of their method is to be taken.
LikeLike
DR. Behe is not an anti-evolutionist. ID is NOT anti-evolution. The debate is all about the mechanisms of evolution.
LikeLike
Oh, piffle! Behe is an evolutionist in the same way most all antievolutionists are, they accept elements of it as OK (microevolutionary variation) but nothing notable. Behe has professed to accept common descent, for example, but has never been caught saying what it is that is commonly descended. It’s a meaningless concession for him, and in his “Edge of Evolution” book he was functionally denying all the class-level origins matters (such as the reptile-mammal transition, which he tried to evade by invoking chloroquine resistance, whose details he muddled, including not addressing the Summers 2014 PNAS paper that Nathan’s Science review noted).
Behe has never laid out what he thinks happened in life, only blips on things he doesn’t like, the dreaded unguided “Darwinism.” Michael Denton is comparable here. someone who allegedly accepts speciation and yet never applies it to anything. That is another major flaw in their position, that they don’t actually have one. And it sounds like you are afflicted with a similar lapse, Frank. Where do YOU think polar bears came from, and when? Explain the data with your view, so we may compare the two to see who can account for the facts better.
LikeLike
Wow, please define “evolution” and then try to demonstrate that ID is against it.
LikeLike
Hi Frank. Or is it Joe, or Sharon, of Frankie, or Virgil?
LikeLike
Where did you get the polar bears from? You do realize that Dr. Behe is just arguing against evolution by means of blind and mindless processes, right? How did you determine blind and mindless processes were responsible for a bear that you cannot account for?
LikeLike
Polar bears originated from brown bears, or did you not pay closer attention to the Liu paper or other systematics on them over the last decade? And what do you think was the cause of polar bears, as your alternative? Or does the non-“blind and mindless processes” model not have to work out what they think happened, or do any of the work to establish it? Perhaps you might not want to be quite so frisky about claiming “cannot account for” in this instance.
LikeLike
Wow, and where did you get those brown bears from? You don’t have a mechanism capable of producing eukaryotes.
LikeLike
Wow you went all “Origins or Bust” in a hurry, Frank. The issue was whether polar bears from other bears, and those on back through carnivora was supported, but you leap at once to the origin of eukaryotes BILLIONS of years earlier. Is that how you avoid thinking about most of the data in between?
LikeLike
No one is arguing that bears are not related. The debate is about the mechanisms involved
LikeLike
Well, that’s a start, so now what is your alternative? Being related to other bears means natural variation in their genomes and subsequent speciation. If you have cause to argue something other than natural or sexual selection is involved (the main toolkit of all evolution, as over a century of work has established), by all means specify them so they may be put to the test. We await your monograph.
LikeLike
The alternative is evolution by means of intelligent design- that being organisms were intelligently designed with the ability to evolve and adapt.
LikeLike
Its clear Frank you have no model, you address no data, you are merely a “not that, no!” sort (to be fair, all ID advocates fall in the same category). Should you ever figure out what you think happened, do let us know, that we may compare your conception to the available technical data, and asses whether your alternative can find favor in the glare of the facts.
LikeLike
“organisms were intelligently designed with the ability to evolve and adapt”
You seem to have invented “deistic evolution”: a god created imperfect replicators, and natural processes took over from there.
LikeLike
I’m not arguing there were perfect replicators at any point, so have no need to suppose any front-loading design to adapt. Organisms just do that, and we can see the nuts and bolts details in the technical literature. Sometimes they run into barriers their natural systems can’t hurdle. We call that extinction. And we’re all now very far either from bear origins or the specific point that Michael Behe tends to cherry pick his science data. Back to point please.
LikeLike
What is the model for evolution by means of blind and mindless processes? How can you test the claim that life arose via blind and mindless processes? You don’t have any supporting data
LikeLike
Back to “Origins or Bust” again, Frank? Please fill us in on what you think went on over the four billion years from the origin of life down to the appearance of polar bears, which it sort of sounds like you do not dispute originated from the adjacent brown bears. So far you’ve shown no familiarity with any of the data, and the prospect of having to reprise the last century of science in an internet chat box seems a fruitless task if none of its likely to sink in anyway. What works have you relied on to have missed the obvious: what we see in living things (and in fossils) is accounted for by natural branching common descent, fueled by natural variations on which natural selection acts. The reptile-mammal transition is one obvious segment of that, but take any subject you care to, please discuss the data, and in what respect you think any of that exceeds the load limit of the aforementioned observed natural processes. If all you have done is channel the Intelligent Design literature, you will be ill-prepared for that task, as they avoid 90% of the data for you.
LikeLike
Hi Joe!
Where did you get the name “Frank Huggins” from? Why didn’t you stick with your female personality “Sharon Mahoney” you tried as PeacefulScience? That one sure worked out well, didn’t it? 🙂
LikeLike
“a mechanism capable of producing”
The basic mechanism of abiogenesis, biological evolution, and all human design work is:
trial-and-error plus memory plus massive parallelism.
Trial and error: Edison tried thousands of materials and constructions for light-bulb filaments before finding one that was practical; Einstein tried several different mathematical functions to base General Relativity on. The ToE divides this term into random variation (trials) and natural selection (success vs. error).
Memory: evolution implies an increase in complexity and efficiency over time; this requires some forms of memory (e.g., genes) to pass initial results on to succeeding generations for further improvements. E.g., the filaments in an incandescent light bulb today are not the first, second, or best (bamboo fiber) that Edison was able to find. Light bulbs evolved, as all human designs have. According to archaeological evidence, it took humans many thousands of years to find the wheel and axle design. Once you have that basic design, it can evolve into gears, pulley, windlasses, water wheels, et cetera.
Massive parallelism: makes unlikely things likely. If one person (only) buys a Power Ball Lottery ticket once a year, the lottery will probably never be won; if millions of people buy a ticket every day, there is bound to be a winner.
There is one more implied ingredient: a universe in which possible solutions exist to be found. If steam engines didn’t work in this universe, no genius would have created one. However if solutions for steam engines and eukaryotes exist, a random search can find them. Why not?
LikeLike
There isn’t a scientific theory of evolution. And ID is OK with evolution by design. What no one can demonstrate is evolution by means of blind and mindless processes actually producing a protein machine. By then way, natural selection isn’t even a search.
LikeLiked by 1 person
@Lino “has been pointed out here, but pointed out by Behe long ago, Summers, et.al., actually confirm Behe’s notion of two amino acid substitutions being needed for resistance to develop.”
Wrong: https://discourse.peacefulscience.org/t/does-summers-et-al-validate-behe/4541
LikeLike
JimV:
Behe’s “The Edge of Evolution” demonstrates, however, that all this “massive parallelism” you’re counting on still isn’t sufficient. In the parasite’s struggle for existence a hundred billion billion replications are needed to simply search the genome for two amino acid substitutions.
LikeLike
His math on that has been thoroughly debunked but that rarely makes a difference to those who have a deep need to believe something
LikeLike
Did you do any source fact checking on Behe’s “Edge of Evolution” argument, Lino? I did (devoting a large section on it in “Evolution Slam Dunk” on the reptile-mammal transition), specifically regarding Behe’s attempt to preclude that macroevolution evidence (none of which he discussed) by using chloroquine-resistance mutations as a supposedly high hurdle nothing significant could cross. Unfortunately he muddled the facts, and the Summer’s PNAS paper “Diverse mutational pathways converge on saturable chloroquine transport via the malaria parasite’s chloroquine resistance transporter” 111 (29 April 2014): E1759-E1767 showed step-by-step the very thing Behe insisted was vanishingly improbable, and Behe has never addressed those data, merely repeating his mantras as if they didn’t exist.
It is perhaps relevant that Behe is a biochemist, someone whose expertise in that field (he did do some work on protein folding decades ago) do not automatically translate into other fields, especially when he doesn’t pay attention to most of the data. And then there are those who read his work through their own filter, taking his arguments at face value without fact checking them.
LikeLike
Nathan:
AS Behe points out, he didn’t do any math in the Eof Evol: he used numbers that have come out of clinical data. These numbers are mere facts, not speculation, which, IMO, adds to the value of EofEvol since it is based not on “theory” but actual case histories.
rjdownard:
Did you also “fact check” his data on the malarial parasite? Isn’t that what the book is about, with extensions to macroevolution simply flowing from what clinical data already is telling us about the “power” of evolution?
LikeLike
Yes I did fact check Behe’s “Edge of Evolution” claims when I did “Evolution Slam Dunk”. I’d cut & paste the whole chapter here but that might be beyond the limits of this venue (Nathan has the text of the book and he may comment on my analysis as he deems suitable). The upshot: Behe started off by confusong mutation rates with rates of fixation, and by avoiding most of the relevant technical literature managed to draw a line in the sand on the improbability of multiple mutations that was empirically refuted by the recent history of chloroquine resistance.
That Behe in turn sought to use this faulty argument to dismiss a priori the reptile-mammal transition data (NONE of which he discussed) said a lot on his tendency to overplay his hand.
Are you claiming that you, Lino, did a source analysis of Behe’s argument? Including reading beyond merely the few sources Behe thought to cite in his argument?
LikeLike
@rjdownard states “[Summers et al] showed step-by-step the very thing Behe insisted was vanishingly improbable”. There are 2 distinct factors in the evolution of chloroquine resistance; a distinction most Darwinists seem unable to grasp. The first is the two initial mutations that confer minimal CQ transport. This is what Behe claims is vanishingly improbable, especially for larger organisms and much, much smaller populations.
The second factor is the step-by-step increase in CQ transport after the initial double mutations to which Behe happily admits can occur with ease. Why is this so hard to understand. Anyone who read The Edge of Evolution can see Summers et al vindicates Behe not refutes him.
By the way, if you read the Summers et al carefully, the authors also state that the mutations have to occur in a specific order. Yet another hurdle to overcome. Easier for malaria; not so much for populations of 10^9.
LikeLike
Kevin, you seem quite good at repeating Behe’s talking points on the Summer’s paper (I made note of his Evo News piece in “Evolution Slam Dunk”), but what you apparently haven’t spotted is that Behe was incorrect in his presumptions. That the mutations couldn’t be that improbable was born out by the incidence of the transitional stages among living populations (a point obvious from the content of the Summers work, which Behe did not address). Unless Behe wanted to suggest that the transitional stages were all specifically designed without having come about by natural mutation from the next (empirically observable) linking population, a degree of micro-managing of design bordering on the perverse, Behe’s gloss on the Summers data is counter-factual.
LikeLike
rjdownard states “That the mutations couldn’t be that improbable was born out by the incidence of the transitional stages among living populations (a point obvious from the content of the Summers work, which Behe did not address).”
Is that so? Then do tell us. Were these transitional stages pre or post K76T and N75E?
LikeLike
Gosh, sounds like you’re again repeating Behe’s argument, but maybe not reading the Summers paper closely. There were several pathways to the finished strain. In “Evolution Slam Dunk” I analyzed one of them, point mutation by point mutation. I will reprise, bearing in mind that ALL of the strains noted are known to exist, no hypotheticals among them:
Starting with D38, the one with that initial N76T mutation you noted, Kevin, the sequence in the variation zone may be represented by the initials “MNTAQNIR”. D38 showed no CQ transport, so that can be our baseline. Any of the billions of parasites on earth might have had that trait for quite awhile, but unless they were exposed to chloroquine, they’d be clearing away the fatal heme just fine, so there’d be no obvious way to tell how long that N76T variant had been kicking around in any ancestral parasite’s DNA.
One mutation in the adjacent 75 spot brings you the D32 strain flips N to E in the second spot, “METAQNIR”, which has a measurable CQ transport rating of 20, meaning some of the chloroquine is getting shunted down the membrane and so not available to interfere with the heme degration going on back in the DV. Unless there were actually chloroquine used on it, though, it’s again unknown when and how often D38s were in play, but selection would now have an edge to work on if chloroquine was used. Incidentally, in the game of genetic shuffleboard, D39 (which also has no initial CQ effect) would have done equally well for a starting point, as “MEKAQNIR” can also flip to the D32 allele in the sequence with only one mutation.
With another mutation at 220, making it “METSQNIR”, you have strain D26, with a CQ rating that surges to 55. You think selection wouldn’t notice that? Each of the next two changes remain at that selectable 55: an alteration at 371 to D20’s “METSQNII”, and one further mutation at 271 resulting in D10’s “METSENII”.
A mutation at 74 produces GB4, “IETSENII”, with a CQ transport value of 100. Selection’s going to be busy then, and parasite merozoites of that strain are definitely going to stand a better chance of making it to the gametocyte springboard stage. Which is confirmed by the fact that GB4 is one of the commoner alleles in the existing Plasmodium population.
Finally, the addition of a change at 326 brings us to the K1 strain, “IETSESII”, fielding an eminently selectable 120 CQ, at the top of the currently known plasmodium haplotypes.
Unless you’re arguing that any one of these existing strains don’t exist, or that it is physically impossible for the single point mutations leading to each to occur (and hence were designed by an specially perverse and sadistic designer hell bent on making life congenial for malaria parasites but hellish for its human victims) perhaps you might consider reevaluating whether Behe’s argument was water-tight.
LikeLike
@rjdownard: your constant, shameless promotion of your self-published book is well noted. However, anyone who bought that book should demand an immediate refund. It’s amusing enough when on every thread there’s that one guy who thinks he’s the smartest one in the room. But when he overplays his hand to the point that he exposes himself as the least, that’s when the fun begins.
I note:
1) Anyone with average intelligence can narrate the path of CQ resistance just by following Fig 3 of Summers et al.
2) Unfortunately, your “close” reading of the paper obviously didn’t tip you off to the fact that the work was done on frog eggs (Xenopus laevis oocytes). Therefore your quaint little narrative is absolutely meaningless. In fact, when tested with minimal CQ transport such as the E2 variant (3 mutations), the malarial cells still didn’t survive in vitro. More on this in #4
3) Your repeated use of N76T (occurs 0 times in the paper) instead of the correct K76T (101 times) demonstrates your true unfamiliarity with the Summers et al paper and seriously calls into question your understanding of the chloroquine debate.
4) rjdownard declares with professorial confidence, “Any of the *billions of parasites on earth might have had that trait for quite awhile, but unless they were exposed to chloroquine, they’d be clearing away the fatal heme just fine, so there’d be no obvious way to tell how long that N76T variant had been kicking around in any ancestral parasite’s DNA.” The literature shows that K76T by itself is strongly deleterious (Lakshmanan et al 2005) so it wouldn’t be “clearing away the fatal heme just fine”.
It’s easy to see why the only publisher who would publish rjdownard’s book is rjdownard.
*Any one of the billion infected victims of malaria has a trillion cells in their body. No one familiar with the literature would use the phrase “billions of parasites on earth”
LikeLike
Thank you for catching my typo, Frank, N should have read K in a few spots in my text, I’ll correct that in future editions. Doesn’t change the point though, and the CQ values are as I stated in the quote. As for the reaction to my book, paleontologist Christine Janis found it most useful (and said so in her Amazon review), so perhaps your negative snark here does not have quite the same impact for me.
LikeLike
rjdownard:
I’m sure you believe everything that you’ve written. I think there’s a little room for criticism; but, you seem to be “straining gnats and swallowing camels.”
Whatever you want to say, the “empirical” facts are that, per White, it takes a hundred billion, billion “multiplications” to produce CQ resistance. As has been pointed out here, but pointed out by Behe long ago, Summers, et.al., actually confirm Behe’s notion of two amino acid substitutions being needed for resistance to develop.
Talk all you want about what Darwinian evolution can develop, but Behe’s results, minus nitpicking, clearly show that Darwinian evolution has a severely limited scope. And, yes, simply does not have the ‘fire power’ to drive reptile-to-mammal evolution.
Your concern about Behe not distinguishing mutation rates from fixation rates is, IMO, irrelevant. Whatever the mechanisms involved in P. falciparum developing CQ resistance, at some point it has to become “fixed” in the population or otherwise it is meaningless because this is a hurdle all populations have to overcome. Let’s not forget that the malarial population size affects ‘fixation.’ In fact, if we multiply the factor of 1 x 10^4 that separates the expected number of replications needed for resistance for both atavouquine and CQ by the average population size of 2 x 10^4 (which comes from a 2013 study), we get 2 x 10^8, which is surely the size of, or, larger than the genome size of the P. falciparum. So, yes, let’s “correct” Behe’s CCC to 10^16 needed replications. Does this, then, now make the reptile-to-mammal transition likely via Darwinian evolution?
I couldn’t possibly agree that it does.
Finally, how is it that you weigh in on Behe’s work, criticizing him for entering the realm of population genetics as a mere biochemist, when, from what I found at Amazon, it appears you’re not degreed in biology? I’m a little puzzled by this.
P.S. I’m not going to “get into the weeds” with you here because I get tired of arguing with people that, in fact, “the sky is blue.” Deny it all you want.
LikeLike
Then by all means explain to us what YOU think was going on with all those therapsids, Lino? If not natural mutation, what? Present your model, account for the data. Was it an unnatural process? If so, tell us, so we may compare your explanation to that of Behe (oh, that’s right, Behe didn’t offer one). Tackle probainognathids in particular, since they were explicitly predicted on evolutionary grounds (a unique double-jaw format unknown in living animals and yet a necessary transitional step to get from the basal reptile to derived mammal layout) and definitely existed.
LikeLike
rjdownard:
Please don’t hyperventilate.
You confuse the fact of evolution with the theory of evolution. One is empirical; the other is theoretical, or speculative.
Thus, you ask me to speculate on what brought about that which we know to be the ‘fact’ of evolution. Nothing wrong here. Except this: the way in which you frame your request is meant to imply that if the Dawinian theory of evolution were abandoned then science would come to a halt. This just isn’t the case.
Physicists have no explanation for the fine-structure constant. They almost don’t even know where to begin to look for the answer. Yet, that doesn’t prevent them from successfully using this constant in their calculations.
Similarly, though we cannot exactly state “how” intelligent design took place, the concept, hypothesis of intelligent design still allows us to ask the right questions about how cellular life operates. Instead of expecting mutations to be entirely random, for example, we should expect them to be biased, or directed. And, in fact, this seems to be the direction current biology is now headed.
It’s not my job to convince you of facts; it’s your job to allow facts to convince you of what they imply.
Behe’s books and papers give ample evidence–as in actual results of experiments and clincial studies, to convince one that Darwin’s theory is simply not powerful enough to explain the evolution of life (which is a fact). But here the death knell of Darwinism is not religion, but genetics.
Darwin, like most scientists of his time, believed an egg to be some kind of goo. We now know much better how complicated the egg cell is with all of its organelles and genetic apparati. He was wrong. He didn’t know better. We do. It’s time to move on.
LikeLike
Both of us read Behe’s works, Lino, but only one of us apparently takes the extra step of fact-checking Behe at source level, which involves of course consulting the relatively few primary sources Behe invokes in his arguments, but also the much broader field of relevant primary source data that Behe does not discuss, and hence does not account for. I know by direct inspection of the Summers paper and others how skewed Behe’s spin has been on chloroquine, and he’s been just as slippery on all of the topics he weighs into, and most glaringly the reptile-mammal transition macroevolution data remain unaccounted for in all antievolution apologetics, not just Behe’s glib hit-and-run effort in “The Edge of Evolution” (I did a full survey of them after all, so don’t have to guess at this omission).
LikeLike
@JimV: You seem to like the example of Edison and his trial-and-error approach to discovery. You used it over on Lenski’s blog as well. Trial-and-error is the result of a certain degree of ignorance on how to obtain a desired result. Edison is always praised for his perseverance
But Nikola Tesla had a different take on this
Your comment on massive parallelism makes the case for ID rather than RM/NS. Enormous populations on the order of trillions and hundreds of millions of trillions like E. coli and p. falciparum respectively, can successfully, randomly search for some solutions whereas larger organisms simply don’t have time on their side.
Last time I checked, steam engines were designed not randomly found.
LikeLike
Whenever a discussion of biological evolution ends up a battle of analogies between Edison and Tesla, you know its straying from the path. Antievoltionists (whether full creationists or the Creationism Lite that is ID) have been monumentally unproductive in all relevant fields for a very long time. Behe is just the latest installment of people motivated by a religious desire to doubt natural evolution, but as close inspection of his books reveal, the level of omission far exceeds the direct data discussion, let alone the accounting of it. Should ID advocates ever graduate to the level where they can account for the available data (tackle that reptile-mammal transition case, for instance, which 100% of them have avoided at data floor level, including Behe), and outperform the evolutionary model in the prediction and practical utility department, let us know. But the deck-stacking 3 Card Monte IDers offer so far carries weight only with people predisposed to agreeing with it.
LikeLike
What would an rjdownard reply be without a shameless shill for his self-published book?
LikeLike
What a remarkably lame and evasive response: attack the messenger. But I did not invent the reptile-mammal transition, though as you snarkily noted, I have written a book about it (of which I am quite proud, and which content qualified for a solid recommendation by Christine Janis, a paleontologist highly primed to have spotted any inaccuracies in it). But my penning a tome on it doesn’t get you off the hook, Kevin: the data still exist, and I observe no antievolutionist has got within a mile of it. So be the first, pioneer the field, Frank: explain why there were probanognathids, and why an evolutionist (Robert Broom) was able to predict their exact nature based entirely on evolutionary principles, decades in advance. If designed, was the Designer especially fond of evolutionists to so contrive a fossil record to please them so? And, while you’re at it, explain why not a single Intelligent Design advocate has even deigned to mention them, let alone account for them. Write a book about it all, if you wish.
In lieu of that, is this how you avoid dealing with contrary data, Frank? If so, it would be all too consistent with how all antievolutionists manage to sustain their convictions in such obvious defiance of the facts, including the present matter of Mr. Behe’s latest book.
LikeLike
My apologies for calling you Frank, Kevin. A slip of the typing. But there still probainignathids.
LikeLike
No less lame and evasive than your response to me the last time you called me Frank in this section. SMH
Since my remarks on Edison were not directed at you, a lame response is all you’re entitled to. You, sir, may have the last word…which I won’t bother reading.
LikeLike
Cool & Simply put….I can put my signature below this!
LikeLike
From Liu, et. al.’s paper:
I wonder what you think of this quote taken directly Liu’s paper.
LikeLike
I’d say it supports the idea that the variants that polar bears have are mostly likely to enhance the gene function in which they occur. Remember, they are talking about the same genes between humans and polar bears, not the same precise mutations.
LikeLiked by 1 person
But the HDiv:Pred of Table S7, as Behe shows in his EN post, show that they are “probably damaging,” while Liu, et. al., go on to detail how the mutations in each of the “nine” genes have been shown to be harmful in other mammals, presumably mice. How is this not support for what Behe has stated?
BTW, I’ve tried to post some of this same basic data at Coyne’s blog, and he simply deleted them. I guess I’m banned there. How nice.
LikeLike
You are missing a very important point. It seems that Behe is missing it too, so please listen to this point very carefully. Let’s focus on apoB. This protein is absolutely key for clearing cholesterol from the blood. Polar bears have a diet that is extremely high in cholesterol (and saturated fats) yet don’t suffer from heart disease. Therefore, they must have enhanced functions somewhere in this system. Yes, mutations in APOB humans have been identified that lead to problems with cholesterol clearance, and cardiovascular and metabolic disease – this shows that APOB is important with that process (something we know through lots of other research also). But the polar bears have different mutations. And they DONT have cardiovascular disease. In fact, they have great protection from it. AND the polar bear mutations are not distributed randomly, but clustered (more than half!) in the domain that is important for clearing cholesterol. So, we don’t know what the mutations do yet (but this is very testable), but the weight of the evidence is strongly in favor of their enhancing apoB, not diminishing it.
Also, and I’ve resisted making a big deal of this because it looks like hand-waving, but the predictive algorithms aren’t very reliable because they don’t take into account the unique context of the protein or the function of the nearby domain. They just analyze amino acid substitutions and make a guess. The imprecision is seen in how often the two algorithms disagree. Yet Behe is basing his entire argument on one of those algorithms at the cost of ignoring everything we know about apoB in other animals. It’s not just a weakly supported position, but one that flies against the face of all the evidence we have. This is very sticky egg on his face and I’m sure he is really regretting using this as his opening example.
LikeLiked by 1 person
Nathan:
This is the nub of the issue: we DON”T know what the mutations do. Behe doesn’t make the claim that Darwinian evolution never increases the fitness of an animal or organism, but rather that the Darwinian process of RM+NS is tilted towards ‘breaking and blunting’ already existing genes, rather than some ‘beneficial’ mutation arising and becoming fixed, is the more likely means of the fitness increase.
But the number of mutations involved certainly is significant. So, I see your point there.
LikeLike
Received my copy of Darwin Devolves today and see that Behe makes his comments on the polar bear in passing–not a detailed look. He says that the mutations are mostly “damaging” per Liu’s Table S7. Nothing new here. We’ve already covered most of this.
But it did afford me a chance to go back and take a closer look at Liu’s paper. I found that although there were 9 mutations in all for APOB, only 4 of those mutations affect the “protein structure”–which I would interpret as meaning these mutations have some significanc affect in how APOB functions. So, for sheer number of mutations involved, for argument’s sake, I think the better number is 4, and not 9.
LikeLike
Seems to me the question is HOW mutations “enhance” function. Sounds like Behe is saying that even mutations that “enhance” function are almost always achieved by damaging a gene with a pre-existing function or turning on or off regulators of that pre-existing gene. If so, then, simply alluding to “enhanced function” doesn’t help destroy Behe’s argument.
LikeLike
Yes it does. Behe is very clear when he says that the mutations are adaptive because they diminish/degrade the function of the gene in which they occur. At least in this example, he’s almost certainly incorrect.
LikeLiked by 1 person
Well, you just said apoB is key for clearing cholesterol, so it already has that function. Enhancing an already existing function is well within the limits of evolution Behe described in The Edge of Evolution. Could it be that whatever mutations polar bears have are like antibiotic resistance where mutant bacteria are missing a transport system and therefore are not affected by the antibiotic?
LikeLike
How can “damaging” be referring to protein stability or binding affinity, but then “beneficial” be used to describe morphological effect? People here are comparing apples to oranges. It doesn’t help to have an almost perfect solution that would work well if it didn’t fall apart or have terrible binding affinity… if it actually falls apart and won’t bind. That’s the whole point of the metric.
LikeLike
since 2012 Polyphen has a possible result of “unknown”. This paper was published in 2014. All of the results are “benign”, “possibly damaging”, or “probably damaging”, with none “unknown”. This is a strong signal. What is the significance of a signal that could have been unknown, but instead is always neutral or damaging?
LikeLike
Ok, lets says that the polar bear’s white coat was caused by “damaged” genetics. Pretty coincidental, because that white coat helps it to survive in the Arctic, but ok, whatever. Does Behe think that other adaptive changes to the genetics of polar bears are also “damaging”? It can swim for many, many miles (it has to since its world melts once a year, but it still needs to eat and get about). The brown bear doesn’t. Coincidence? Its elongated neck so as to see better swimming in water. Coincidence? Its heavily matted and webbed feet that help it in traction on the ice and to swim in comparison to the brown bear’s bare feet. Coincidence? Its smaller ears to reduce the loss of heat. Coincidence? Its sunglass-like eyelids. Coincidence? Its more carnivorous teeth and other adaptations to an all-meat diet (the brown bear is an omnivore by contrast). Coincidence? It’s large stomach capacity to enable opportunistic feeding. Coincidence? Its 4″ layer of fat that helps it stay much warmer in the arctic then the brown bear could. Coincidence? Their ability to fast for NINE MONTHS while recycling their urea. Coincidence? I guess If you say so, Behe. All “damaging” mutations.
LikeLike
By the way, you can read the above info from https://en.wikipedia.org/wiki/Evidence_of_common_descent#Vertebrates_3
LikeLike